Cyprinodontiformes vivíparos e ovovivíparos
Livebearer Cyprinodontiformes
Espécies > Ficha da espécie > Xiphophorus hellerii
Species > Species profile > Xiphophorus hellerii
|
|
|
Cyprinodontiformes vivíparos e ovovivíparos Livebearer Cyprinodontiformes Espécies > Ficha da espécie > Xiphophorus hellerii Species > Species profile > Xiphophorus hellerii |
||||||||||||||
|
|
|
||||||||||||||||||||||||||||
|
Uso de imagens : Todas as imagens deste sítio não podem ser usadas sem a devida permissão. Os direitos de autor de cada uma das imagens pertencem ao respectivo fotógrafo/autor que as tirou/criou e não podem ser utilizadas sem a respectiva autorização. A
fim de de poder usar as imagens de acordo com os respectivos direitos de
autor/licenciamento/legislação aplicável e condições deste sítio,
por favor envie-nos uma mensagem de correio elctrónico.
Estão disponíveis para consulta mais imagens sobre esta espécie na galeria de imagens.
Limite de tolerância : 13ºC e 35ºC Limite de sobrevivência : 9ºC a 11ºC e 39ºC a 41ºC ( dependendo das populações ) pH ideal : 7 – 8.2 dH ideal : 9º - 19º Salinidade máxima : 1,011 ( 15,4 ppm )
Já vai sendo inclusivamente vulgar encontrar, incorporado no respectivo material genético destes indivíduos, vestígios de outras espécies do género, o que resulta de cruzamentos ocorridos algures num passado recente em cativeiro, nomeadamente com as espécies Xiphophorus variatus e Xiphophorus maculatus. Entre as populações que se encontram na natureza existe já por si uma certa variabilidade, a qual é quase impossível de testemunhar nos animais cultivados. Para se ter uma ideia, ainda que muito superficial, consulte-se na Galeria a página relativa às variedades selvagens de Xiphophorus hellerii. No final da década de 1960 considerava-se mesmo que existiam várias subespécies de Cauda-de-Espada :
Xiphophorus hellerii strigatus, ( Regan ) : México - Rio Papaloapan e Rio Coatzacoalcos Xiphophorus hellerii guentheri, (Jordan & Evermann ) : México - Laguna Sontecomapan, Rio Grijalva, Bacia hidrográfica do Rio Usumacinta ( México e Guatemala ); Guatemala e Belize – rios que desaguam no Atlântico, e em toda a área da distribuição geográfica no território das Honduras. Xiphophorus hellerii signum ( Kallman & Atz ) : Guatemala - Rio Chajmaic. Xiphophorus helleri alvarezi ( Rosen ) : México e Guatemala - Rio Lacantún.
Xiphophorus helleri alvarezi, Rosen, 1960; ( valido como ) Xiphophorus alvarezi, Wischnath & Foerster, 1985
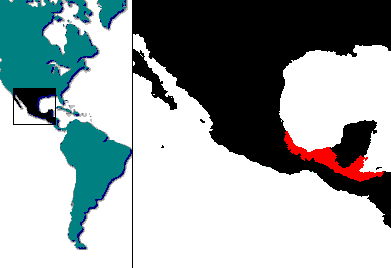
Devido à sua distribuição geográfica relativamente ampla, encontraram-se mesmo diferenças interessantes em termos biológicos e morfológicos entre populações distintas. A selecção sexual desempenha neste âmbito um papel preponderante. É notória a preferência das fêmeas por machos que exibam a espada caudal mais longa, e coloração mais atraente, ( Jerald B. Johnson & Alexandra L. Basolo, 2003 ). O problema reside no facto de que os machos mais coloridos serem também mais facilmente vistos e capturados pelos predadores. O facto de uma fêmea se deixar cortejar por um macho demasiado fora dos padrões mais harmoniosos com o ambiente que os rodeia, pode inclusivamente colocar também a sua própria vida em risco, dado que passam os dois a constituir um alvo fácil para os predadores, ( Jerald B. Johnson & Alexandra L. Basolo, 2003 ). Para além do apêndice caudal e da coloração propriamente dita, há outras marcas que são comuns nos animais selvagens. Alice Marrin Kerrigan, no seu trabalho “ The Inheritance of the Crescent and Win Spot Marking in Xiphophorus hellerii “, descreve pormenorizadamente o papel da genética na transmissão de algumas destas marcas. Tal como acontece noutras espécies de Poeciliídeos, ocorrem por vezes dois tipos diferentes de machos dentro do mesmo cardume. No caso do Xiphophorus hellerii, esta situação foi descrita nomeadamente em relação à população do Rio Jalapa. Num dos dois tipos, os machos caracterizam-se por possuírem a linha característica ao logo do corpo de cor escura, ( “ machos de linha negra “ ), enquanto os restantes apresentam a linha lateral vermelha ou acastanhada ( “ machos de linha vermelha “ ). Mesmo atingindo a maturidade sexual antes dos machos de linha vermelha, os machos de linha escura nunca se tornarão dominantes no grupo. Mesmo quando ainda são 3 a 5 mm maiores do que os seus oponentes da linhagem vermelha são habitualmente subjugados por estes nas disputas. De qualquer forma, os machos de linha negra são sempre mais pequenos do que os seus concorrentes ( quase metade do tamanho ), porque esses continuam a crescer até atingirem a maturidade sexual, a qual ocorre só daí a muitas semanas, ou mesmo meses mais tarde. A preferência notória das fêmeas pela linhagem vermelha e a sua dominação sobre a linhagem negra não levou aquela à extinção daquela. Os machos da linhagem vermelha transportam genes que permitem perpetuar o tipo concorrente, para além do facto de que os da linhagem negra poderem acasalar mais cedo, ainda que contanto com eventuais menores taxas de sucesso no acasalamento. Outra diferença notável, para além de factos comportamentais distintos, é que as crias descendentes da linha negra são maioritariamente do sexo masculino, enquanto as da linha vermelha são fêmeas, ( Dierk Franck, Agnes Müller & Natascha Rogmann, 2003 ). De toda a forma, o sucesso no acasalamento por parte dos machos não depende exclusivamente da sua fisionomia estar mais ou menos de acordo com as preferências do sexo oposto. Esta espécies caracteriza-se por comportamentos violentos de dominação, sobretudo entre os machos, os quais resultam em competição pelas fêmeas e geram violência. Tais conflitos podem mesmo ser observados em aquário, ainda que a amplitude destes desafios e os seus resultados dependam não só da herança genética dos peixes, mas igualmente das dimensões do aquário, da composição do grupo e da concentração de indivíduos. O tamanho dos exemplares nem sempre é sinónimo de sucesso nestas contendas. O aspecto fisiológico e a motivação podem mesmo resultar da submissão de um indivíduo maior perante um menor. Entre grupos desta espécie há sempre uma complexa rede de relações e forma-se mesmo uma hierarquia, comportamento esse que é observável mesmo em aquário. Por essa razão convém ter em consideração que o espaço vital para a manutenção saudável de um grupo deve ter sempre reflectir 3 factores muito importantes, designadamente que os animais adultos podem chegar aos 15 cm, que os animais dominantes perseguem os restantes, ( sendo que daí podem resultar mesmo ferimentos mortais se os perseguidos não tiverem espaço para se refugiarem ) e, por último, que esta espécie possui comportamentos sociais relativamente complexos. Tais comportamentos são abordados com uma interessante profundidade no artigo “ Social organization of small heterosexual groups of Green swordtails ( Xiphophorus helleri, Pisces, Poeciliidae ) under conditions of captivity “, ( Jacques P. Beaugrand, Jean Caron & Louise Comeau, 1984 ). Tal como acontece com outras espécies do género Xiphophorus, e não só, têm surgido ao longo do tempo inúmeros testemunhos de reversão sexual ( o indivíduo pertence funcionalmente a um dos sexos mas geneticamente pertence ao oposto, ou “ muda de sexo “ a partir de certa altura ). Ainda que não seja muito vulgar, este fenómeno acontece mesmo em populações criadas em cativeiro cujas origens são desconhecidas. Essas linhagens incluem habitualmente no seu material genético os vestígios de cruzamentos entre variedades e populações diferentes, pelo que o seu valor como matéria de estudo científico é questionável. Ao contrário do que se acredita, não basta manter no mesmo aquário apenas fêmeas para que uma delas se transforme em macho, ou vice-versa. O que é interessante é verificar que as fêmeas que deram à luz várias ninhadas de crias e se transformam em machos podem fertilizar outras fêmeas e produzir descendência. Uma experiência relativa à reversão sexual completa foi muito bem documentada no artigo “ Complete Sex-Reversal in the Viviparous Telost Xiphophorus helleri “ ( J. M. Essemberg, 1926 ). Na natureza estes peixes são omnívoros, alimentando-se de quase todos os organismos vivos de dimensões convenientes, nomeadamente zoobentos e zooplâncton, invertebrados aquáticos, crustáceos aquáticos, insectos e suas larvas, mas igualmente de bastante matéria vegetal variada ( sobretudo detritos vegetais e algas filamentosas ou unicelulares ). A alimentação usual em cativeiro não apresenta problemas. O Xiphophorus hellerii tolera muito bem alimentos industriais secos ( flocos ), frescos ou congelados e dá resultados espectaculares quando recebe também alimentos vivos. Uma dieta adequada corresponde às dos outros ovovivíparos mais comuns, sem especiais cuidados a assinalar. Para além de alimentos liofilizados em flocos, recomenda-se vivamente artémia salina, larvas de mosquito, tubifex, dáfnias, todos estes vivos ou congelados. Como complemento aceitam bem a carne moída de moluscos ou da maior parte do peixe de mar usado na alimentação humana. Muito raramente também se pode aproveitar a carne crua sem gordura e também moída de animais domésticos, ( vaca, porco, aves e coelho ). Existe uma diferença assinalável em termos de crescimento e fecundidade entre os peixes alimentados frequentemente com alimentos naturais e aqueles que só têm acesso a alternativas artificiais ( D.P. Kruger, P.J. Britz & J. Sales, 2001 ). Esta é uma espécie de reprodução contínua, desde que seja mantida dentro dos limites de temperatura e foto-período ideais. Em cativeiro, o canibalismo de recém nascidos é frequente e muito eficaz. A quantidade de recém nascidos que podem ser eliminadas pela própria mãe situa-se na ordem dos 32,5% dos nascidos, sendo que 85,2% dessas vítimas foram apanhados pela fêmea no fundo e 14,8% a superfície, de acordo com um estudo realizado em laboratório ( C.L.W. Jones et al, 1998 ). Nesta mesma investigação é sugerido que, para se salvar o maior número de crias, devem ser previstos suficientes abrigos à superfície. No entanto, esta é uma espécie consideravelmente prolífica. A fêmea Cauda-de-Espada apresenta um período entre partos que varia normalmente entre 24 e 30 dias, sendo muito frequentes os ciclos de 28 dias. Esta duração está intimamente relacionada com a temperatura e com a alimentação. A duração de um parto pode ir de sensivelmente 2 horas a mais de 10, ( C.L.W. Jones et al, 1998 ). O intervalo entre os primeiros partos pode ser irregular e acontecer entre os 27 e os 90 dias. A fertilização ocorre cerca de uma semana após um parto. Uma cópula com sucesso pode dar origem a 5 a 9 gestações, dado que as fêmeas podem conservar no seu corpo esperma viável por um período que, nesta espécie em particular, pode chegar aos 2 anos. Nas regiões tropicais a reprodução pode ser contínua. A temperatura ideal para a gestação situa-se entre os 22ºC e os 26ºC, sendo suspensa qualquer actividade reprodutiva abaixo dos 15ºC e acima dos 29ºC. As fêmeas sujeitas a angústia ou colocadas em gaiolas do tipo “ maternidade “ poderão recusar-se a dar à luz por períodos longos. Independentemente da duração da gestação nessas condições em particular é comum uma parte considerável de crias daí resultantes nascerem mortas ou com problemas graves. Um parto pode dar origem de 20 a 40 crias nas fêmeas mais jovens e até pouco mais de 240 nas plenamente desenvolvidas. O número de crias das fêmeas idosas e o intervalo entre os partos tendem a replicar de novo o início do ciclo reprodutivo. A alimentação dos recém nascidos segue os parâmetros habituais dos ovovivíparos da sub-família Poeciliinae.
A destruição do meio
ambiente através de contaminação com águas residuais domésticas e
industriais, captações de água acima da capacidade de reposição pela
precipitação para fins agrícolas, para fins industriais ou para consumo
humano, a par da introdução de espécies exóticas, são as maiores ameaças
que o Xiphophorus hellerii enfrenta actualmente. Para esclarecimento de dúvidas e mais informações consulte-se Cyprinodontiformes vivíparos e ovovivíparos no Livro Vermelho de Espécies Ameaçadas da UICN.
|
© 2006 - 2011 Todos os Direitos Reservados e-mail : viviparos.com
Enquanto os conteúdos são assim considerados, algumas páginas contêm imagens e material sujeitos a direitos de autor e usados apenas sob permissão. Tal informação está apenas disponível para uso privado, ensino ou investigação. Devem ser obtidas permissões dos detentores dos direitos de autor para outros usos. Por favor examine bem caso por caso e assegure-se das respectivas autorizações sempre que necessário.
A comunicação e conteúdos são transmitidos no pressuposto de que não há garantias absolutas da sua exactidão e incontestável perfeição. Os visitantes e utilizadores deste sítio são convidados a ponderarem sobre o carácter provisório de alguns dados antes de os utilizarem, pelo que se assume da responsabilidade desses utentes o tipo de uso que lhes é dado. A comunicação e conteúdos são transmitidos no pressuposto de que não há garantias absolutas da sua exactidão e incontestável perfeição. Os visitantes e utilizadores deste sítio são convidados a ponderarem sobre o carácter provisório de alguns dados antes de os utilizarem, pelo que se assume da responsabilidade desses utentes o tipo de uso que lhes é dado. |